§ 1. THE PROBLEM OF GROWTH. An immediate approach to the laws of human nature may be made through the problem of growth. Why does an organism grow? The answer to this question solves the basic problem for centuries inherent in the concepts of urge and instinct; it makes intelligible the evidently spontaneous character of behaviour, the searching, hunting, purposive, voluntary aspects; it reveals the secret of acquisitiveness and inventiveness; it removes the riddle of the new increments that are constantly emerging in the course of the learning process; it squares the idealistic side of human life with the facts of physiology, neurology and physics. It is the bridge over which the laws of dynamics are carried from the realm of the physicist to the realm of the psychologist, where they become the laws of human nature.
§ 2. GROWTH IN PHYSICAL WORLD. There is growth in the physical world. Systems of energy form and disintegrate. Chemical evolution is a slow growth process. Growth is occurring at a more rapid rate in biological evolution with a technique different in form but not in principle. In the growth of living organisms an evolution of startling speed is taking place with still a different technique, but again the same in principle. The enormous differences in speed between cosmic, biological and individual growth, trace down to the differentiation of energy systems into more and more heterogeneous and unstable configurations of energy. In the large it is a picture of cosmic life shifting phenomenologically from inorganic to organic forms.
Growth is a puzzle to the physicist who measures the life of an energy system by its dissipation of energy into that most unruly and chaotic form of all, namely, heat. If this is the total picture, the universe is running down. But what about growth? Is it achieved at the expense of more energy than is saved to the emerging compound and the growing organism? Is it the cosmic plan that organic and intelligent life evolve under the law of diminishing returns? Is Nature’s great experiment a failure? Is it to happen that growth, achieved at the expense of more energy than is saved, will result in intelligent systems of energy, capable of turning upon the heat lost in their production and converting it into useful forms? Does growth, after all, mean a winding up process that “saves”, as it goes along, more heat than is lost to organisation? Or do cosmic rays represent the gain in our immediate system that compensates for its losses? Such questions might sound foolish were it not true that sooner or later the growth process as a whole must be envisaged in terms of dynamics, and that the facts of cosmic and biological evolution must square, on the one hand, with physics and, on the other, with ethics. There is no getting around the point. If man’s behaviour reveals a forward reference, if it is hunting, searching, purposive in its character, as observation proves it to be, these characteristics have analogues in all other energy systems. Ethical man, with his philosophy of values, a developing, thinking being, operating according to one set of principles, could not survive or even articulate with a cosmos based upon an incompatible set of principles. Man must go to the cosmos, or the cosmos must come to man; the one process is the same as the other.
§ 3. COMPARISON OF GROWTH AND GRAVITATION. If a falling body in a gravitational system is an intelligible phenomenon, so is growth, because the two are functionally similar. The gravitational system furnishes the stress under which the apple hangs on the tree; it gives to the apple its possibility of motion. At the same time it gives to the apple the remote end of its fall, and the direction of its fall. The system contains the apple’s past and its future; and both, simultaneously, are conditions of the present, the momentary behaviour of the apple. The fall, as a given continuum, can be accounted for only by going outside that continuum to the system of conditions that sustain it. If man were the apple he would, let us suppose, be in dynamic relation to the centre of the earth, but without knowing it. He could not see into the future.
Growth is a movement. Like any movement, as a finite problem, it has its beginning in a potential. Let us call it in this case a growth potential. The potential is derived from a cosmic system operating in accordance with the laws of dynamics; that system is at the same time furnishing the line of growth and its remote end or goal. The growth process is the realisation of that potential in kinetic energy, or movement, when the balance of the system is disturbed, just as the falling of the apple is the realisation of gravitational stress. What, in growth, corresponds to the breaking of the apple’s stem? Stimulation of the living organism by its environment. What is the system? It is the world of energy in which we grow; the cosmos of life in which, as a race, we are evolving. Wherever there is movement, the suppositions of a potential and a remote end are not only legitimate but necessary; for otherwise movement is a meaningless occurrence having no beginning, no direction and no end.
Growth is something observed. We wish to make it intelligible. We do so by postulating the growth potential and by defining the growth process as the resolution of that potential toward a remote end. The remote end for the individual organism is natural death. The end for organic evolution is a problem in philosophy. The end that religion postulates as a personal entrance into immortality is neither refuted nor substantiated by known facts; for this sort of immortality and the identity of the immortal are not experimentally demonstrable. In general, however, organismic logic demands some sort of immortality consistent with man’s place in the cosmic plan, and an immortality which will harmonise with the laws underlying existence. But again man’s place in the cosmic plan, and an immortality consistent with it, are philosophical questions.
§ 4. RESPONSE TO FUTURE. According to the law of configuration, the beginning, the direction and the end of a given movement are aspects of the same descriptive unit. Each is significant only with reference to all the others. Growth, then, in terms of dynamics, is a fiction, something undeserving of a name, unless it be regarded as a response to the future. The realising of any potential is a response of one object or another to its future. It is in this sense that the behaviour of the apple contains the functional analogues of spontaneity, searching, hunting, purpose.
Potential is a guarantee of action. It is convenient to say that the potential demands its own resolution. It shares this demand with the whole of which it is a part, for the whole gives it the potential. It is therefore legitimate to say that the organism demands growth once the growth potential is established. It is already set up in the living organism. If the organism cannot grow it dies. Growth is a total life movement, an expanding, differentiating pattern of motion exhibiting itself, on the one hand, in an enlargement of structure, and on the other, in behaviour. But for growth to take place, a potential must be maintained and released by environment. This is accomplished in several ways, all reducible to one, namely, the activating influence of fields of force surrounding the organism—oxygen, heat, pressure, light. To these forces must be added the energy assimilated through food. So much for the growth that exhibits itself in the enlargement and differentiation of organic structure. This type of growth merges over gradually, with no break, into the other, the development of behaviour, which also must be sustained and released by environmental forces. The process is then called stimulation. The complex organism is so differentiated in structure that these sustaining and releasing forces act upon different systems of organs, oxygen through the lungs, other chemicals through the digestive system, and stimulation through the nervous system. But all this is the same in principle.
§ 5. PURPOSE. If, then, behaviour, in the large, is a product of a growth potential we can understand how it must be creative, inventive, and self-expressive. These, in fact, are its major characteristics. It is essentially emergent. Moreover, each act of behaviour demands completion in the attainment of a goal, subordinate to the remote ends of growth.
More immediately, behaviour derives its purposeful and seeking character, on the one hand, from neural tensions, and on the other hand, from those environmental situations to which the tensions are related. Anything that is external to the nervous system may be regarded as environmental; for it is the nervous system, primarily, with which are concerned the tensions that behaviour resolves. Part of this environment consists of organs within the body such as glands, the digestive tract and reproductive organs. These as well as external forces, furnish stimuli that induce nervous tensions. But this internal form of stimulation, so important in the behaviour of complex organisms, has been encountered, before, in principle. A gravitational system is continuously subject to internal disturbances that release potential into kinetic energy. The stem of the apple breaks and lets the apple fall. The breaking of the stem is analogous to intraorganic stimulation in the living organism. The breaking stem is something external to the apple and its line of motion toward the centre of the earth. Muscle contractions of the stomach wall that stimulate the nervous system are, similarly, external to the nervous system and to the behaviour of seeking food. Both the hunger response and the apple falling are movements toward remote ends, the one a food goal, the other, the centre of the earth. Theoretically each motion ceases in the same way, for the same reason; the apple when, on its way to the centre of the earth, an obstruction balances its own potential against it; the hunger response, when the food goal is reached and the stomach filled. Both motions cease when an unbalanced system of energy returns to a condition of equilibrium.
§ 6. EXAMPLES. Here is a scientist in his laboratory absorbed in the intricate processes of chemistry, creating a new theory and testing it one way and another; hunting, searching, calculating, and finally arriving at a great discovery. Months of arduous and painstaking endeavour were required; the problem beckoned with a call both persistent and irresistible; the labour was gratuitous; the goal was its own reward. But again, in principle, all this has been encountered before. From the bowels of the earth, under pressure, a stream of water comes to the surface as a spring. The nervous energy of the scientist boils into fruition, so to speak, under the pressure of a maturation potential. It is released in a behaviour called inventiveness. A fountain of water in the one case; in the other a fountain of knowledge. The spring continues to flow; a surface stream is formed. Tirelessly, irresistibly, persistently, winding, tumbling, falling, dammed up and overflowing some obstacle, on it goes, down, perhaps, hundreds of miles, requiring months of time, until a water-level is reached. Through the maze of contours offered by mountains and plains it goes, never stopping until the end. What is the source of this behaviour, of this persistence? It is a gravitational system giving direction, unerring, in the line of least action. A human being, alive, planning, thinking, under his own type of gravitational tension, in this case a maturation potential, guides his ideas, the muscles that speak his words and the hand that wields his pen, winding, tumbling, held by an obstacle of difficult logic only to overcome it, delayed by an intricacy of mechanical technique only to discover the solution, until the remote end, the discovery, the reaching of neural level, and resolution. Through the maze of contours offered by the limitations of his knowledge which his own system provided him, just as the gravitational system provides its own mountains and plains through which the stream must find its way, the chemist directed his stream of thought over an elaborate route, yet in the line of least action.
Why does the chemist’s behaviour seem voluntary while the stream seems to flow “involuntarily”? Only because, as men, we are behind our goals, moving toward them. When we think of the stream we see the whole situation, the source, the stream, the water-level; the picture is complete; the whole that is conditioning its parts is evident. But let man imagine himself the stream at any particular point, unmindful of the remote end that corresponds to the chemical discovery, and the whole that conditions its parts is now not evident. The stream is “searching”; its behaviour is “voluntary”; it is going on because it “wants” to; its “want” is the tension under which it flows; it is “voluntary” because the remote end is in the future.
It is not mind that gives to the chemist’s performance its voluntariness except in a phenomenological sense; will involves no new cosmic principle. Voluntariness must be defined dynamically in terms of ends not reached. Phenomenal mind exists apart from voluntariness in this sense. A movement of man is voluntary in virtue of being conditioned by a whole, whose field property may be described by the phenomenological term “will”. It is the act that is voluntary, not the will. So too the stream is “voluntary” in the sense of being conditioned by a gravitational system; the system itself is not. Voluntariness is a characteristic of the act, not of the actor. Common usage of terms has long recognised this fact. The will is free, not voluntary. If we consider the stream as an isolated thing, its force is equivalent to voluntary effort; the influence of the gravitational system upon it is equivalent to will. Man has will when considered a relatively segregated whole, operating under his own potential; the effort he expends, in moving, accrues to muscular contraction. The contraction corresponds to the stream. The reason why will is a term applicable only to man and animal as a mental phenomenon, lies in the difference of form into which the energy-system, man or animal, is structured, as compared with the form into which the earth, streams, mountains and plains are structured. Phenomenologically, man has a will and the earth has not. Dynamically, as potential, the will of man and the dynamics of a gravitational system are similar. Energy is the will of Nature, anywhere, in the sense that, whatever its manifestations or structurisations, the same laws and the same logic apply to it.
§ 7. OBJECTIONS MET. But, you say, do not inform us that man in his longings for truth, in his loves and idealisms, is no more than a gravitational system. Take care! It is easy to fall back into the dualistic and mechanistic trap. What is idealism? As a fact of experience it belongs to man. It differentiates man from systems of gravitation like the earth; it differentiates man from the beast. Phenomenologically then, to be idealistic is to be human, not to be an animal, nor to be inorganic. But, in terms of dynamics what is idealism? The setting up of remote ends for which to strive. Man does it himself relative to a cosmic plan. So too a simple gravitational system sets up ends that must be “striven for” in a dynamic sense. In accordance with its relation to the universe, the system furnishes its own centre toward which the apple hangs under stress; the system breaks the stem of the apple; the system supplies the fountain and the stream. Herein are the analogues of initiative and spontaneity. Any system, as a unified whole, establishes the ends of the activities that go on within it. That man in dynamic relation to his environment can set up goals, is, in terms of dynamics, nothing new in Nature. His consciousness of a goal is akin to the apple’s stress. Dynamically there is no inorganic versus organic; no man versus beast; no mind versus matter; no life versus inert bodies; no plan versus chaos; no teleology versus mechanism; no foresight versus blindness; no initiative versus passivity. Any whole—an atom, man, it makes no difference—contains all the principles pertaining to reality; it is in itself a universe. The cosmos may be likened to a fraction. Energy is the single denominator; phenomena of all forms are the numerator. The laws of dynamics belong to the former; the levels of complexity and degrees of purposiveness to the latter.
§ 8. SUMMARY. We have attempted to show, at some length, the consequences of the growth potential. They are the voluntary, hunting, inventive characteristics of man’s behaviour, characteristics that accrue to the demanding nature of unresolved tensions. Temporary searchings are expressions of a larger, more general potential “behind” them, a potential that is not only “behind” but “around” them in space and time; for it exists with reference to a point far in the future; it is the growth potential seen as part of a cosmic plan. Man spends his life searching, willing and resolving tensions. Out of the growth potential emerge, through differentiation, the broader aims of life; to live, to acquire knowledge, to contribute to posterity, and to believe in the future. The more detailed aspects of behaviour are further differentiations from the same whole, occurring, like the local windings and tumblings of the stream, in the presence of temporary obstacles in the course of daily life. The continuance of a performance always signifies a goal ahead. All events in Nature involve a forward reference.
We are now ready to examine the facts of development in a typical animal embryo. From these facts it will become evident that behaviour is an outcome of growth and is not to be explained by the usual, mechanistic hypotheses.
§ 9. EVIDENCE FROM NEUROLOGY: DEVELOPMENT OF MOTILITY IN THE EMBRYO. The nervous system of the embryo develops as an expanding, differentiating unit. At first it is a total, relatively homogeneous pattern of cells. Local and specialised nerve centres, or groups of cells, emerge and assume relative, not complete independence. Throughout the growth of the nervous system, field properties condition the activities of specialised parts. One would expect, therefore, to find illustrations of organismic principles in the development of the embryo’s motility; and this is precisely what we do find.
The first movements of the embryo, taking the salamander studied by Coghill1 as an example, are those of the organism-as-a-whole. These are waves of contraction, called flexion, that pass down the body of the animal from the head region tailward, curling the animal into a semicircle or perhaps a complete circle. A little later a wave down one side of the body will not have been completed before a subsequent wave starts down the opposite side, curling the animal in the opposite direction. This results in an “S” shaped coil. When this movement takes place with the proper frequency it propels the animal in a primitive fashion through the water. When the forelegs develop, which precedes the growth of the hind legs, there is no separate nerve supply that permits local movement. The legs move only as the body moves; the movements are aspects of trunk movement. This is necessitated by the fact that nerve fibres to the trunk branch and carry impulses to the forelegs. The same situation holds also for the hind legs. Subsequently nerve centres that supply the legs differentiate sufficiently from the total nerve pattern to permit local reflexes when the legs are stimulated. Formerly it was thought that these local reflexes combined to form complicated activities like swimming and walking; but this is not the case. These local reflexes will not integrate. Coordinated movements must wait upon further development of the total nervous system of the animal. Rhythmic and properly timed movements of all four legs emerge together, already integrated, from the expanding and differentiating total nerve pattern. Thus the walking movements are not a product of reflexes nor of local random movements that combine as a consequence of practice or experience. Co-ordinated movements result from mautration. Reflexes are end products of differentiation, not units of integration.
Perhaps more striking yet is the manner in which separate movements of the leg develop. When the leg acquires relative independence of movement, it is the leg-as-a-whole that moves. Gradually, as the nerve supply maturates, movements of the fore part of the leg emerge, but this segment at first moves as a whole. Then movements of the “hand”, or foot-as-a-whole, differentiate from the forearm; and, finally, movements of the fingers or toes differentiate from hand-movements-as-a-whole. In each instance a part of the body, relatively segregable from the body as a whole, functions as a totality; and, in turn, part activities of that particular member emerge, integrated with pre-existing wholes as they emerge. Involved in all this differentiation is the organism-as-a-whole. The differentiation never leaves mass action behind.
The most interesting of all developments in the motility of the salamander occur in the anterior region of the body. Before the animal is able to walk with all four legs it can lift the anterior part of its body with its front legs, moving at once the head, neck, shoulders and legs. This it will do in response to a food object, say a fly, before movements of the head and jaws can be made. The animal lunges toward the fly before it can eat! Subsequently head and neck movements emerge from the total anterior body movements; jaw movements, in turn, emerge from head and neck movements. At this period in the animal’s growth, if a fly is brought near its head it will actually seize and crunch the fly in its jaws. But this is before swallowing movements can be made! No better illustration could be found of the principle that the organism’s growth is a unified process, taking place under its own potential with reference to a remote end and without experience on the part of the animal. The forward reference of the movements just described can be accounted for in no other way. The future, as well as the past, controls the present.
That the salamander should snatch a fly before it can eat is no more remarkable in principle than the simple fact that before an apple can fall the last part of the distance to the ground it must traverse the first part. The first part looks forward to the last in the same sense that catching a fly before eating it is possible looks forward to the later period of maturation when eating is possible. The maturation process is a continuum, just like the fall of the apple, conditioned by a whole external to it. The embryo’s movements are kinetic realisations of maturation. Their developmental phases correspond exactly, in terms of dynamics, to the temporal phases of the apple’s fall. Both the fall and the growth process are units in time as well as in space. In its growth, the salamander is merely obeying the laws of least action and configuration. The concept of instinct is superfluous.
The situation in the case of the salamander is harder to comprehend only because one cannot put his finger, so to speak, upon the remote end of the growth process so readily as he can put his finger upon the centre of the earth in the case of the falling apple. It is easy to see how the fall must be a continuum conditioned in its totality by an alignment of stresses. It is easy to see how one stage of the apple’s fall will not account for the next; although the one is necessary for the other. The one is necessary for the other because both are necessary, by definition, before there is anything to explain. Units, conditioned as wholes by larger wholes, are the only kinds of explicable phenomena. This is the law of determined action. Any other situation yields indeterminism. And nothing is a unit unless it is a complex whole, a differentiated field with respect to space and a continuum with respect to time.
Another way of regarding the forward look of the apple’s movement through the first foot of space, let us say, is to reflect that the movement (no obstacle interfering) through the last foot of space is guaranteed by the system before the first phase takes place. In the case of the salamander, lunging toward a fly before snatching is possible, and snatching before eating is possible, seem like intelligent anticipation. But the embryo need not know that at a later stage in its development it will swallow the fly and enjoy it, any more than the apple must know that at last it will rest upon the ground. The continuum of nervous and mental development is a species of movement continuum occurring in a cosmic system of energy surrounding the organism both in time and space. A cosmic plan guarantees the end of growth as the gravitational system guarantees the apple’s flight.
§ 10. FIRST MOVEMENTS OF THE INFANT. Studies of the human fœtus unmistakably bring out the same principles, demonstrable not only in the salamander but in the toadfish, rat and rabbit. Turning now to the newborn infant, studies by Irwin2 point definitely to a continuance of the same growth precedure. The first co-ordinated movements are of the body as a whole. The apparently random waving of arms and legs and squirming of the body are not discrete, isolated reflexes occurring in chaotic fashion, but are mass actions variable in appearance because the infant’s skeleton and muscular structure are anatomically so complex. Each apparently separate movement is an aspect of a total body movement, only part of which is detected by the observer. The infant cries at first only as the whole body moves; and its first attempts voluntarily to control its hands and feet are made together with contraction of the whole body. It is true, however, that as embryos develop the trunk plays, overtly, a less dominant rôle. Likewise, in the infant’s later movements the rôle played by the trunk may at times be only that of assuming a general tenseness with no overt action, while the extremities are the parts exhibiting the greatest motility. Certain local movements emerge more discretely than others from the total pattern. Nevertheless, it is evident that parts of the body not actually moved still function in the total pattern. The importance of total body co-ordination even in adult performances can be demonstrated in all kinds of situations. Try, for instance, to aim a rifle with the body in an awkward position; try driving golf balls in an awkward stance.
§ 11. MATURATION. From all this it becomes obvious that the development of motor co-ordination is a function of maturation, not of practice or experience. This principle is of profound significance for the problem of learning in children and adults, and is rather damaging to the orthodox belief that “practice makes perfect”.
§ 12. ALL MOVEMENT DETERMINED BY END. A given muscular movement obtains its direction as does the falling body from a system of stresses, in this case from a system of neuromuscular tensions. Each movement is made with reference to a remote end. The remote end, in the case of complex movements, is not, as in the case of the gravitational system, a single point, but a situation more akin to the remote end of expansion of gas in a container, or the remote end of radiation phenomena. This situation, in the moving organism, involves what, for purposes of convenience only, may be designated as outer and inner aspects of the remote end. Consider a hungry person approaching the cupboard for something to eat. The neuromuscular system is under tension, in this case set up and released in large measure by intraorganic stimulation from the stomach wall. The person is nervous, irritable and tense. The remote end of his movements, so far as the organism as such is concerned, is within the organism itself; a relaxed, equilibrated state of the neuromuscular system. But the movements that end in relaxation are movements that bring the person to the cupboard and result in a filled stomach. The resolving process carries the organism to the external aspect of the goal-situation, the food-object. Thus, reaching the outer and inner aspects of the goal-situation are but two ways of describing the same process. Organic relaxation and arrival at the external object are coincident.
The direction of these movements is conditioned by a total situation involving arrangements of external stimuli toward which the organism is always in dynamic relation. We must think of the moving organism as an object moving in a larger dynamic whole, much as we think of the apple and its line of flight. There is, seemingly, a difference between an apple and the organism in that the apple does not propel itself toward the goal while the animal or human being does. This difference is only relative, however, and requires no new principles of explanation. The difference demands only a more complicated application of these principles.
The apple has mass, and does, in a sense, propel itself toward the ground, for as a mass, defined in terms of the whole, it makes a contribution to the whole. It possesses an increment of whole-character. To the extent that it is part-of-a-whole it is self-propelling in terms of the law of determined action. The living organism exhibits definite whole-properties, in its own right, that overtly figure in the process of reaching the goal. It directs its own movements toward the goal to that extent, and exercises the same government over its parts that the whole of which it is a part exercises over it. While the picture in the case of the organism is more complicated, phenomenologically, it is no more complicated in principle; the difference in the two situations is handled by repeated applications of the same set of laws as we go from phenomenologically more simple to more complex situations. At the same time that an organism, moving under stress in an environment, is being conditioned by a larger dynamic whole which furnishes at once the stress, the remote end, and direction of movement, so, in terms of the same set of laws, the organism gives direction, as a system itself, to the movements of its own parts.
§ 13. GOALS. In this complicated picture the contribution made by the organism results in alternative goals within the general goal-situation; that is, there are several points in the environment of which the organism is a part that may correspond to the single point in the centre of a gravitational system. The goal for the hunger response may, for example, be the cupboard, or the refrigerator, or a box of candy on a table in another room. The organism lives and moves, therefore, in an environment to which it is so adjusted that the larger whole amounts to a great number of related gravitational systems in one, each system being a particular alignment of stresses in the organism dynamically related to different stimulus-situations, and to each other.
§ 14. APPLICATION TO ETHICS. Were we to apply this line of reasoning to problems in ethics we would attempt to show, in detail, how intelligence figures in a human situation in a way similar to mass in a gravitational system. Suppose, for example, that a small object falls a given distance upon a glass plate. No damage will be done to the plate. But let a large object fall upon the plate from the same distance and the plate will be smashed. This is our cue to the rôle played by the part in the whole, although under the laws of the whole. The system, and the object in it, share “responsibility” for the broken plate. The larger object broke the plate because of its greater mass and therefore greater potential for performing work. Society and the individual share in the ethical responsibility for the deeds of man; society because it is the whole from which man derives his moral standards, and the individual, because he is the immediate agent of the deed, executed in terms of his judgement. His judgement corresponds to the mass of the falling body. The welfare of another individual, or of society as a whole, corresponds to the plate. Society itself corresponds to the gravitational field.
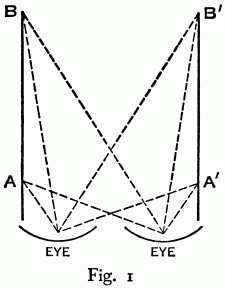
§ 15. DYNAMICS OF NEURO-MUSCULAR SYSTEMS. Returning to the main thread of discussion, there are certain neuro-muscular systems in the human organism, simple enough to permit a fairly clear conception of their dynamics. The optic system is one, as Koffka has so admirably pointed out. Here are two eyes each supplied with three pairs of muscles. These muscles all contract and relax as a unit as soon as the optic system is sufficiently mature. No practice is required.
Let it be assumed that the eyes of the infant are at rest, focussed upon a candle at A. The candle is moved to B. Both eyes follow the candle. But suppose that instead of starting at the point A we start at A′. The infant’s eyes would follow the candle quite as readily from A′ to B′. Notice, however, that the situation is quite different. In the first case muscles on the left side of each eye are relaxed, and contracted on the right; in the second case they are contracted on the right and relaxed on the left. In the two cases the light falls on different parts of the retina. Consequently, the neurological structures more actively concerned in the two movements are different in the two cases. In fact they are different for every position from which the eyes commence to move, for every different direction of movement, and for every different extent of movement. Indeed, it is impossible to conceive of any two movements just alike.
We are in a hopeless situation if we accept the experience theory. An infinite number of preformed pathways and bonds would have to be presupposed. If we try to account for these movements in terms of practice, or in terms of inherited reflex pathways where bonds are assumed between a stimulus and a given movement, one would never be able to make eye-movements; for each is different from every other with respect to its mode of stimulation through the retina, and its stimulation through tension and relaxation of the separate muscles. (The muscles themselves contain sense organs whose stimulation by muscular contraction or relaxation aids in the production of movement.) Obviously there must be an explanation that accounts for eye-movements on an entirely different basis, ruling out practice and the concept of reflex action.
§ 16. CONSTRUCTION OF RETINA. Note how the retina is constructed. The spot of clearest vision is in the centre, the fovea. There is a gradient of sensitivity that passes from the fovea to the margin of the retina. Whenever the brightest spot in the field of vision does not fall upon the spot of greatest sensitivity on the retina the optic system is disequilibrated. Tensions are set up in the eye-movement system. The equilibration is effected when the spot that is brightest in the visual field falls directly upon the fovea. The gradient of sensitivity across the retina and its dynamic relation to the optic system condition the simultaneous coordination of six pairs of muscles. If the eyes move so that the stimulus falls on another part, no closer to the fovea, there is no resolution of tension; if the eyes move so that the stimulus falls farther out on the retina there is an increase of tension. Only by moving the eyes so that the object falls upon the fovea is the tension resolved. Thus, eye-movements follow the law of least action just as the falling apple does. In the case just cited, and in all others, the law of least action and its corollaries permit of no explanatory concepts in psychology such as those of reflex action and the acquiring of motor co-ordinations by means of practice.
§ 17. EXPLANATION BY GROWTH-POTENTIAL. The explanation here offered is traced back to the growth-potential. The optic system is an expanding, differentiating whole, capable of functioning “perfectly” upon reaching a sufficient degree of differentiation. The differentiation is maturation. Reflect that the laws of movements in living organisms are the same as the laws of falling bodies. An apple need not try by experiment to reach the ground; its fall is not a selection of possibilities from a host of random performances, nor does it matter whether the apple is in the United States or in China. It has never fallen before; it does so perfectly the first time. From the standpoint of science there are no perfect or imperfect movements, not even in human conduct. In terms of dynamics Nature makes no errors and requires no practice. There are no wasted movements; there is no trial and error.
§ 18. RAT AND GUINEA PIG. Consider the difference between a newly-born guinea pig and a newly-born rat. The former can walk quite readily at birth; the latter is relatively helpless. The difference is one of maturity, not practice. The rat that must “learn” how to walk is merely demonstrating maturation in an environment of objects as his co-ordinations improve; while the guinea pig matures in the uterus. In both cases maturation is a function of stimulation, not of exercise. What seems to be exercise in the rat, and consequently an explanation of learning, is only the growth potential released into kinetic energy by environmental instead of uterine stimuli. Learning depends on practice in the same way that the falling apple depends on movement. The two are the same. To attempt an explanation of the one by the other is only to set up a fictitious problem whose solution is as impossible as the problem is fictitious.
§ 19. DIFFERENTIATION AND MATURATION. There have been infants who, artificially prevented from walking until long after the natural period of “trying”, walk perfectly well in the first attempt. (Perfectly must always be used in a practical, not a scientific sense.) Birds reared in high nests fly successfully the first time they try. Experiments on frog and salamander embryos by Carmichael3 show that these creatures may be anæsthetised and prevented from making swimming movements over a period of days, while unmolested embryos are passing through the first awkward stages of swimming. When the drug is removed, the anæsthetised animals swim quite as well as the normal who have had several days of “practice”. The awkwardness of the latter in the interim consists of those grosser, less differentiated movements that any living organism makes in early stages of maturation. Differentiation and maturation go hand in hand.
Professor Bird,4 experimenting upon chicks, discovered that pecking movements improve without practice within the limits of time the chicks can live without food. While practised chicks performed better than the unpractised, there was no evidence that faster progress in the latter was caused by experience. They had food, were stronger, maturated faster, and had the advantage of light; while the unpractised chicks were kept in the dark. Other experiments show that, without practice, forcibly-fed chicks will improve in pecking with age, until the limit of about two weeks is reached, when they no longer will peck at all. They will then die in the presence of food. Here we have evidence that maturation will take place only within limits in the absence of stimulation (Padilla).
1 Cƒ. Coghill; Anatomy and the Problem of Behaviour; New York, 1929.
2 Cƒ. Irwin; The Amount and Nature of Activities of Newborn Infants, etc.; Gen. Psy. Monog.; 1930; Vol. 8; 1-92.
3 Cƒ. Carmichael; Development of Behaviour in Vertebrates Experimentally Removed from the Influence of External Stimulation; Psy. Rev.; Vol 33; 1926; 51-58; Vol. 35; 1928; 253-260.
4 Cƒ. Bird; The Effect of Maturation upon the Pecking Instinct of Chicks; Ped. Sem., Vol. 33; 1926, 212-234.
| PREV | NEXT |
| Table of Contents |
| For Kindle e-readers: humannature.mobi |
| For EPUB e-readers: humannature.epub |
| Back to | THE SCIENCE OF COMPREHENSION: How We Understand and Reason About Abstract Concepts : The New 21st-Century Paradigm Shift | |
| Back to | Deley’s Homepage |